Ein Hauptproblem bei der Verarbeitung sensorischer Informationen ist das sogenannte 'binding', d.h. herauszufinden, welche ankommenden Informationen zusammengehören, also ein und dasselbe Objekt beschreiben. Im Visuellen läuft dies oft auf die Erkennung von Grenzen zwischen Objekten oder den Kanten der Objekte hinaus. Ein Artgenosse zum Beispiel könnte eine von der Vegetation im Hintergrund völlig verschiedene Farbe aufweisen. Da visuelle Objekte meist räumlich zusammenhängen, ist die Strategie, Grenzen zwischen ihnen zu bestimmen um sie einzeln zu isolieren oft ausreichend.
Auch in der auditorischen Verarbeitung stellt sich die gleiche Frage, aber in mancher Hinsicht ist sie hier schwieriger zu beantworten, denn es gilt die Frage zu beantworten: "Was ist die Begrenzung eines auditorischen Objekts?" In der visuellen Verarbeitung führt eine räumliche Trennung von Objekten direkt zu einer räumlich-getrennten Antwort auf der Retina. Im Vergleich dazu besitzt das auditorische System nur zwei räumlich getrennte Inputkanäle (die Ohren auf beiden Seiten des Kopfes), anstelle von Millionen Retinazellen. Bevor ein auditorisches Abbild der Umgebung erzeugt werden kann, sind einige Verarbeitungsschritte notwendig. Wir konzentrieren uns hier auf zwei Projekte: die Monaurale Echo Unterdrückung und die Erkennung periodischer Signale.
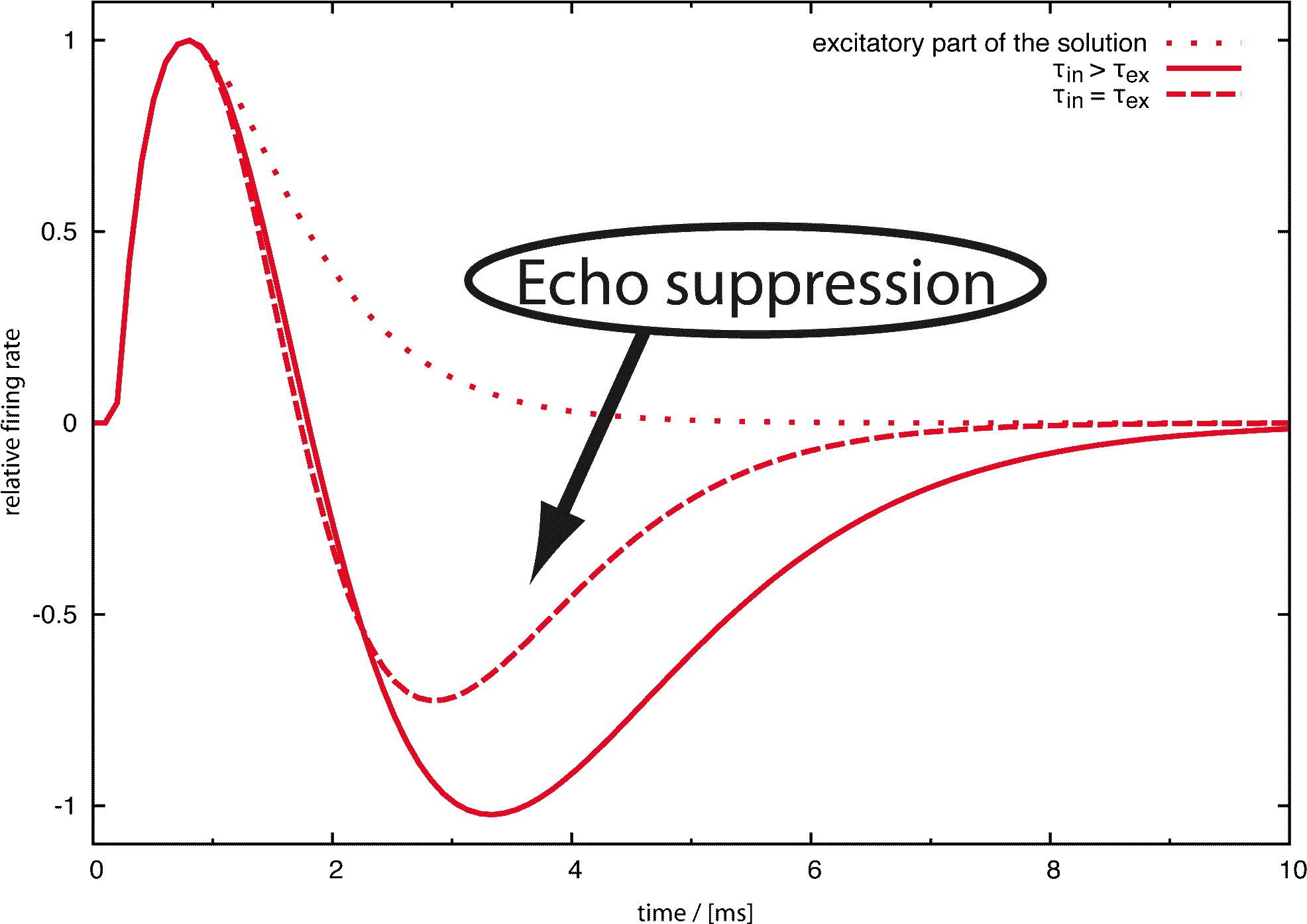
Jedes biologische System, das akustische Signale verarbeitet, muss Störungen des originalen Signals auf Grund von Reflektionen (Echos) bewältigen können. Deswegen ist ein neuronaler Mechanismus, der die störenden Informationen der Echos unterdrückt, oder sogar nutzt, sehr nützlich für die auditorische Wahrnehmung.
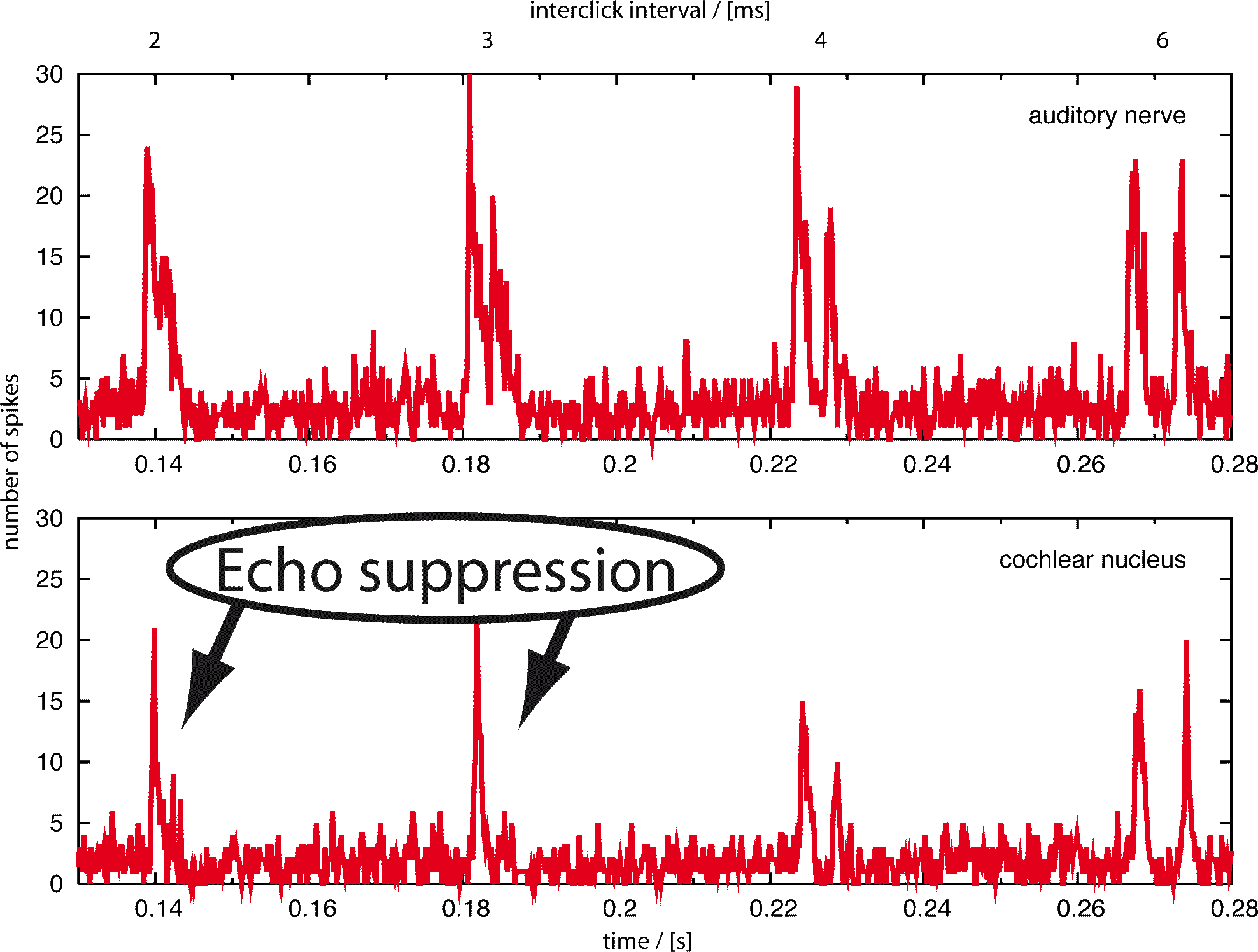
Als Folge werden Echos, die weniger als 20 ms nach dem originalen Signal ankommen, für gewöhnlich nicht wahrgenommen. Ein einfaches Experiment kann zeigen, dass die Echounterdrückung zum Teil binaural und zum Teil monaural ist. In einem großen Raum, zum Beispiel einem Hörsaal, können die sonst unterdrückten Echos leicht durch Zuhalten eines Ohres wahrgenommen werden. Da die Echos in einem kleinen Raum sehr viel schneller sind, funktioniert dieses Experiment dort nicht. Deswegen besteht die Echounterdrückung aus einem langsameren binauralen Mechanismus, der beide Ohren benötigt und einem schnellen monauralen, welcher nur Informationen von einem Ohr nutzt. Seit 1963 (Harris et al.) wurde vermutet, dass der Nukleus Cochlearis, der erste Knoten nach der Cochlea und dem auditorischen Nerv, an der Echounterdrückung beteiligt ist. Das Ziel unserer Forschung war, eine quantitative Antwort auf die Frage zu geben, inwieweit inhibierende Schleifen, die 1990 (Wickesberg and Oertel) im Nukleus Cochlearis gefunden wurden, für das Phänomen der monauralen Echounterdrückung verantwortlich sein können.
Das Modell
Entsprechend den biologischen Tatsachen besteht das Modell aus drei verschiedenen Neuronenpopulationen: Die Neuronen des auditorischen Nervs (AN), die des dorsalen (DCN) die des anteroventralen (AVCN) Bereichs des Nukleus Cochlearis. Um das Modell so einfach wie möglich zu halten, haben wir die gleichen Parameter für die Neuronen des AVCN und des DCN benutzt. Die Zellparameter unserer generischen Neuronen stammen aus der Arbeit von Wickesberg und Oertel, die hauptsächlich "bushy cells" studierten. Die Verbindungen zwischen den unterschiedlichen Populationen wurden ebenfalls der Biologie entsprechend modelliert. Jedes Neuron des auditorischen Nervs ist exzitatorisch mit einem Neuron des DCN und einem des AVCN verbunden. Währendessen erhält das Neuron im AVCN inhibitorischen Input von der Nervenzelle des DCN. Drei Neuronen, die, wie oben beschrieben, verbunden sind, formen eine so genannte neuronale Triade, die Input von nur einem Frequenzkanal erhält.
Im ersten Schritt wurden die Neuronen als Poisson-Neuronen, einem einfachen analytisch beschreibbaren Neuronmodel, untersucht. Im zweiten Schritt wurden Spike-Response Neuronen, eine Verallgemeinerung der Leaky-Integrate-and-Fire Neuronen, benutzt.
Wichtig ist festzuhalten, dass das Modellverhalten bei Variation der Neuronenparameter im Wesentlichen unverändert bleibt. Die von uns benutzten Parameter stammen aus der Publikation von Wickesberg, Oertel und Wu, wobei auch bei anderen Parametersätzen das Verhalten des Models stabil blieb. Die Robustheit unserer Resultate zeigt, dass diese neuronale Verschaltung, welche von Wickesberg und Oertel entdeckt wurde, per se die Eigenschaften besitzt, die viele Autoren dem Nukleus Cochlearis zuschreiben. Danach kann man, wie auch schon Wickesberg und Oertel vorgeschlagen hatten, tatsächlich annehmen dass monaurale Echounterdrückung zumindest eine der Funktionen des Nukleus Cochlearis ist. Ein schnell auf einen anderen folgender Klick wird unter natürlichen Umständen meist ein Echo sein. In diesem Sinne kann der Nukleus Cochlearis als Ort der monauralen Echounterdrückung angesehen werden.
Es ist bekannt, dass Echos, welche 2-20 ms nach dem originalen Signal auftreten, im zentralen Nervensystem unterdrückt werden. Ein Modell der binauralen Echounterdrückung, das Echos bis zu 20 ms unterdrückt, wurde kürzlich vorgestellt. Dieses Modell hat eine minimale Unterdrückungszeit von 5 ms. Somit kann das hier vorgestellte Modell mit einer Unterdrückungszeit von 2-4 ms das Bild vervollständigen.
Eine wichtige akustische Eigenschaft ist die periodische Modulation eines Signals. Somit ist die Erkennung und Identifikation solcher Fluktuationen sehr bedeutend in der auditorischen Verarbeitung. Es ist seit langem bekannt, dass komplexe Geräusche, die aus vielen unterschiedlichen Frequenzen zusammengesetzt sind, dennoch mit einer eindeutigen Tonhöhe (Pitch) wahrgenommen werden. Diese Tonhöhe korrespondiert selten mit einer der Frequenzkomponenten des Signals, als vielmehr mit der zeitlichen Struktur des Signals, der Einhüllenden. Solch eine zeitliche Struktur spielt nach derzeitiger Meinung eine entscheidende Rolle in der Objekterkennung und Identifikation.
Da die zeitliche Struktur des Inputsignals nicht durch die Cochlea erschlossen werden kann, muss ein neuronaler Mechanismus existieren, der Periodizitäten im Signal erkennt. Obwohl schon mehrere neuronale Modelle zur Lösung dieses Problems vorgeschlagen wurden, fehlte bis jetzt eine detaillierte mathematische Studie hierzu. Kürzlich haben wir zur Periodizitätsdetektion ein einfaches Modell, das auf einem Netzwerk von neuronalen Verzögerungsleitungen basiert, analysiert. Wir waren damit in der Lage einerseits die Funktionsweise dieser Netzwerke zu verstehen und andererseits die essentiellen Parameter zu bestimmen, welche die Performance solcher Netzwerke limitieren.
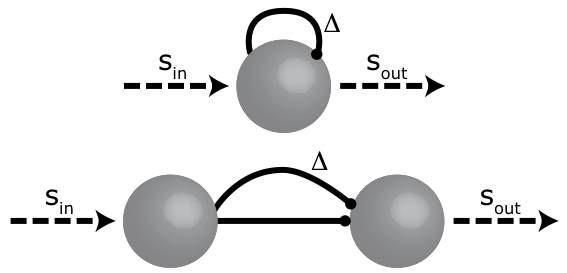 Die der Detektion von periodischen Signalen zu Grunde liegende Netzwerkstruktur besteht aus einer Sammlung von neuronalen Verzögerungsleitungen. Wie in der Abbildung zu sehen ist, kann solch eine Verzögerungsleitung durch eine Feedback-Verbindung oder durch eine Feedforward-Verbindung implementiert werden. Jede Gruppe von Neuronen, die auf diese Art verbunden sind, hat eine bestimmte Verzögerung (Delta), die grob mit der inversen Periode (1/T) der Periodizität korrespondiert. Wenn zum Beispiel ein Signal mit einer Frequenz von 100 Hz amplitudenmoduliert ist, pulsiert das Inputsignal Sin 100-mal in der Sekunde. Wenn die Länge der Verzögerungsleitung 10 ms beträgt, also genau zum Input passt, wird der Output verstärkt, da sich der direkte Input und der verzögerte Input genau überlagern. Wenn die Länge der Verzögerungsleitung nicht mit der Periodizität des Signals zusammenpasst, wird der Output abgeschwächt.
Die der Detektion von periodischen Signalen zu Grunde liegende Netzwerkstruktur besteht aus einer Sammlung von neuronalen Verzögerungsleitungen. Wie in der Abbildung zu sehen ist, kann solch eine Verzögerungsleitung durch eine Feedback-Verbindung oder durch eine Feedforward-Verbindung implementiert werden. Jede Gruppe von Neuronen, die auf diese Art verbunden sind, hat eine bestimmte Verzögerung (Delta), die grob mit der inversen Periode (1/T) der Periodizität korrespondiert. Wenn zum Beispiel ein Signal mit einer Frequenz von 100 Hz amplitudenmoduliert ist, pulsiert das Inputsignal Sin 100-mal in der Sekunde. Wenn die Länge der Verzögerungsleitung 10 ms beträgt, also genau zum Input passt, wird der Output verstärkt, da sich der direkte Input und der verzögerte Input genau überlagern. Wenn die Länge der Verzögerungsleitung nicht mit der Periodizität des Signals zusammenpasst, wird der Output abgeschwächt.
Eine Sammlung von solchen Verzögerungsleitungen, mit unterschiedlichen Laufzeiten, kann ein Inputsignal analysieren und dessen periodische Komponenten herausfiltern. Wir haben gezeigt, dass die zeitliche Genauigkeit der Signale bewahrt wird. Dies bedeutet, wenn einmal ein Objekt durch das Verzögerungsleitungssystem identifiziert ist, ist es möglich das Signal ohne Verlust an Genauigkeit weiterzuverarbeiten. Insbesondere ist die Lokalisation eines auditorischen Objekts mittels interauraler Zeitdifferenz (ITD) möglich.
Es wurde oft vorgeschlagen, auditorische Objekte derart zu identifizieren, dass aller auditorischer Input aus derselben räumlichen Position zusammengenommen werden sollte. Ein Problem dieses Ansatzes ist, dass bei mehreren Schallquellen diese interferieren, was eine Lokalisation der Quellen äußerst schwierig macht. In unserm Modell wird zuerst ein Objekt identifiziert, bevor es lokalisiert wird. Da die zeitliche Präzision nicht beeinträchtig wird, ist Objektlokalisation immer noch möglich, nachdem Aufmerksamkeit zuerst auf ein einzelnes Objekt gelegt wird und somit störende Geräusche von anderen Schallquellen unterdrückt werden.
- Bürck M., van Hemmen J.L. Modeling the cochlear nucleus: A site for monaural echo suppression? J. Acoust. Soc. Am. 122:2226-2235 (2007)
- Friedel P., Bürck M., van Hemmen J.L. Neuronal identification of acoustic signal periodicity Biol. Cybern. 97:247-260 (2007)
- Harris G.G., Flanagan J.L., Watson B.J. Binaural interaction of a click with a click pair J. Acoust. Soc. Am. 35:672-678 (1963)
- Wickesberg R.E., Oertel D., Delayed, frequency- specific inhibition in the cochlear nuclei of mice: A mechanism for monaural echo suppression J. Neurosc. 10:762-1768 (1990)
- Zahn T.P., Neural achitecture for echosuppression during sound source localization based on spiking neural cell models Ph.D. thesis, Technische Universität Ilmenau (2003).
- Pecka M., Zahn T.P., Saunier-Rebori B., Siveke I., Felmy F., Wiegrebe L., Klug A., Pollak G.D., Grothe B. Inhibiting the inhibition: a neuronal network for sound localization in reverberant environments J. Neurosci. 27:1782-1790 (2007)