|
Due to their small interaural distance, lizards have developed a special hearing mechanism, the "pressure-gradient receiver" which is only partly understood. Their auditory peripheral system differs from the one found in mammals by a coupling of the ear drums through internal cavities in the head.
|
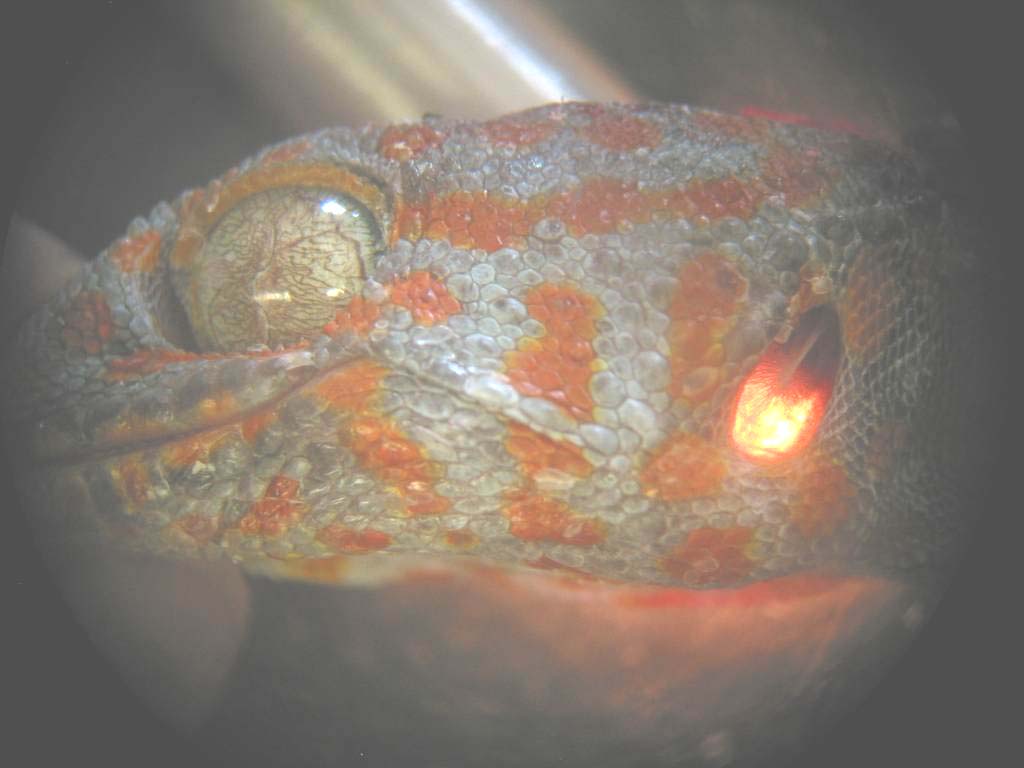
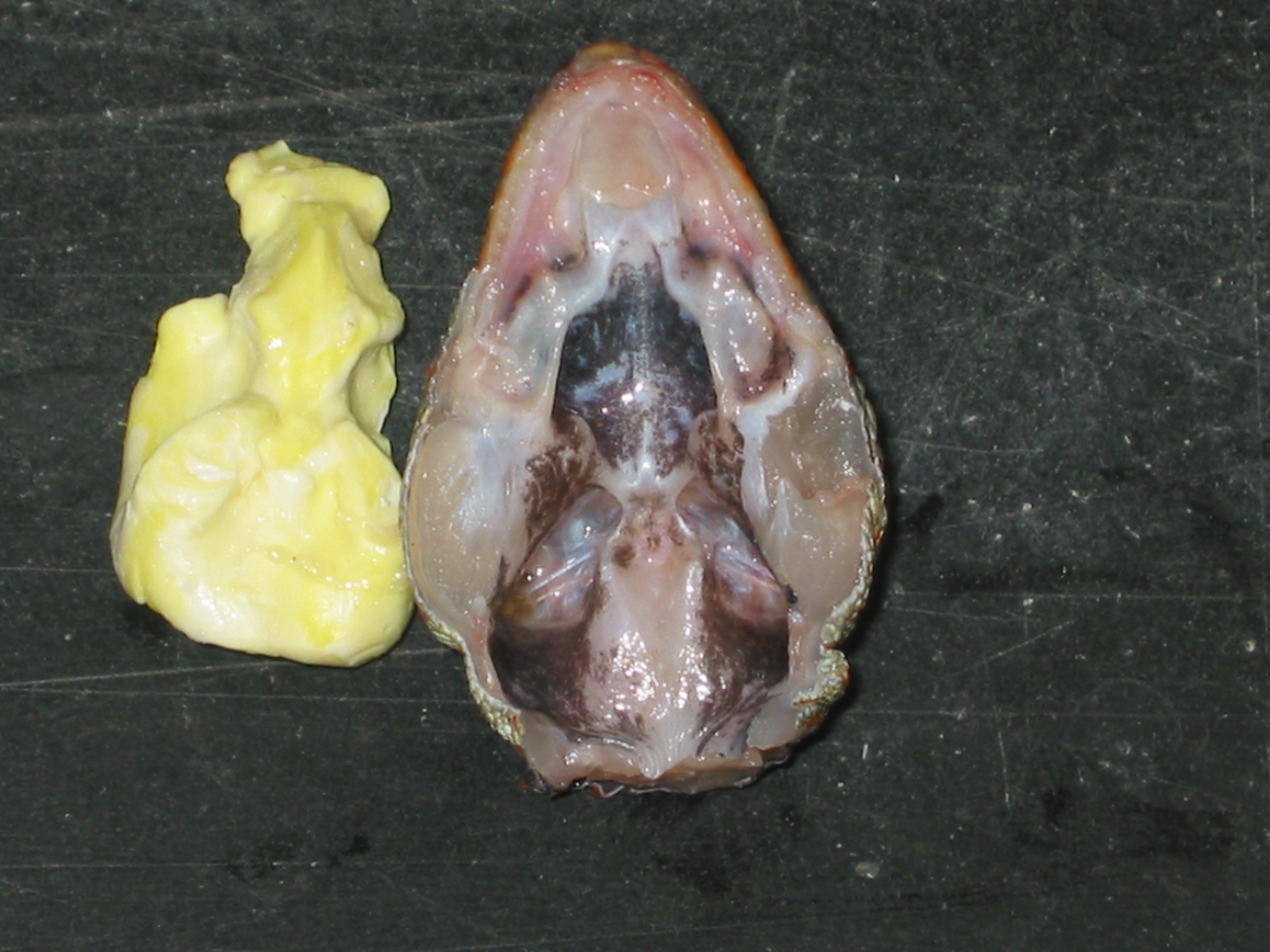
Left: The coupling of the tympani can be visualized easily. If we illuminate the head, especially the tympanic membrane, at one side, you can observe the reflection of the light at the other side.
Middle: Indirectly illuminated tympanic membrane with the ossicle, the so called columella.
Right: Imprint of the internal head cavities.
(Photos taken from Christensen-Dalsgaard)
|
|
The auditory signal processing contains several interesting aspects. Firstly, the ossicle, for the lizard a simple lever construction called columella, is asymmetrically attached to the tympanic membrane. That leads to the question how the middle ear can influence the profile and the frequency of the vibration of the tympanic membrane.
Amplitude measurements of tympanic vibration have been made experimentally as shown in the figure (Measurements from Manley 1977).
|
|
In order to calculate the reaction of the tympanic membranes to external excitation we have to take into account the internal coupling of the tympani. How does this connection influence the differences between the phase and amplitude of the vibration?
Fletcher (reference below) and Christensen-Dalsgaard et al. (reference below) developed an impedance model treating the auditory system as an electrical circuit. In this case, the vibration of the membrane can be represented as combination of the external pressure and the internal vibration fields. The obvious question is if such a model is sufficiently exact. And, if the model works, what influence the internal cavities have, e.g. in generating resonance effects.
|
|
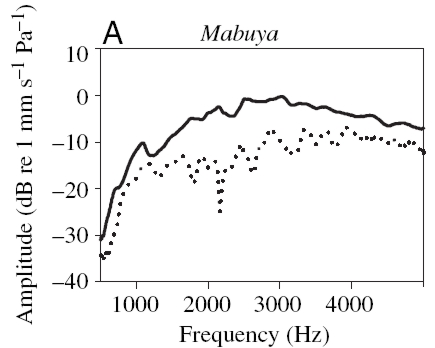
Vibration amplitude of the membrane as a function of frequency with ipsi- (solid line) and contralateral (dotted line) input. (Measurements taken from Christensen-Dalsgaard 2006)
|
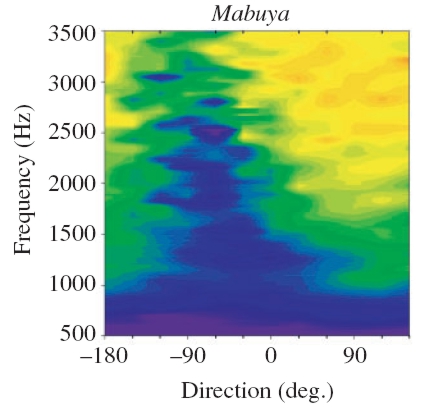 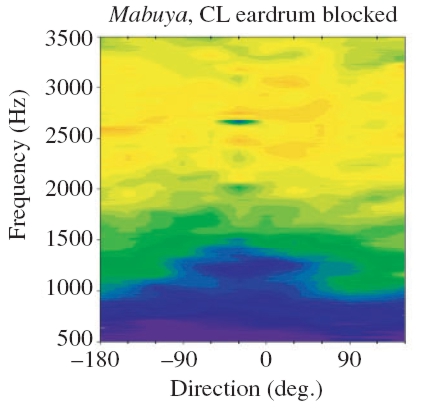
|
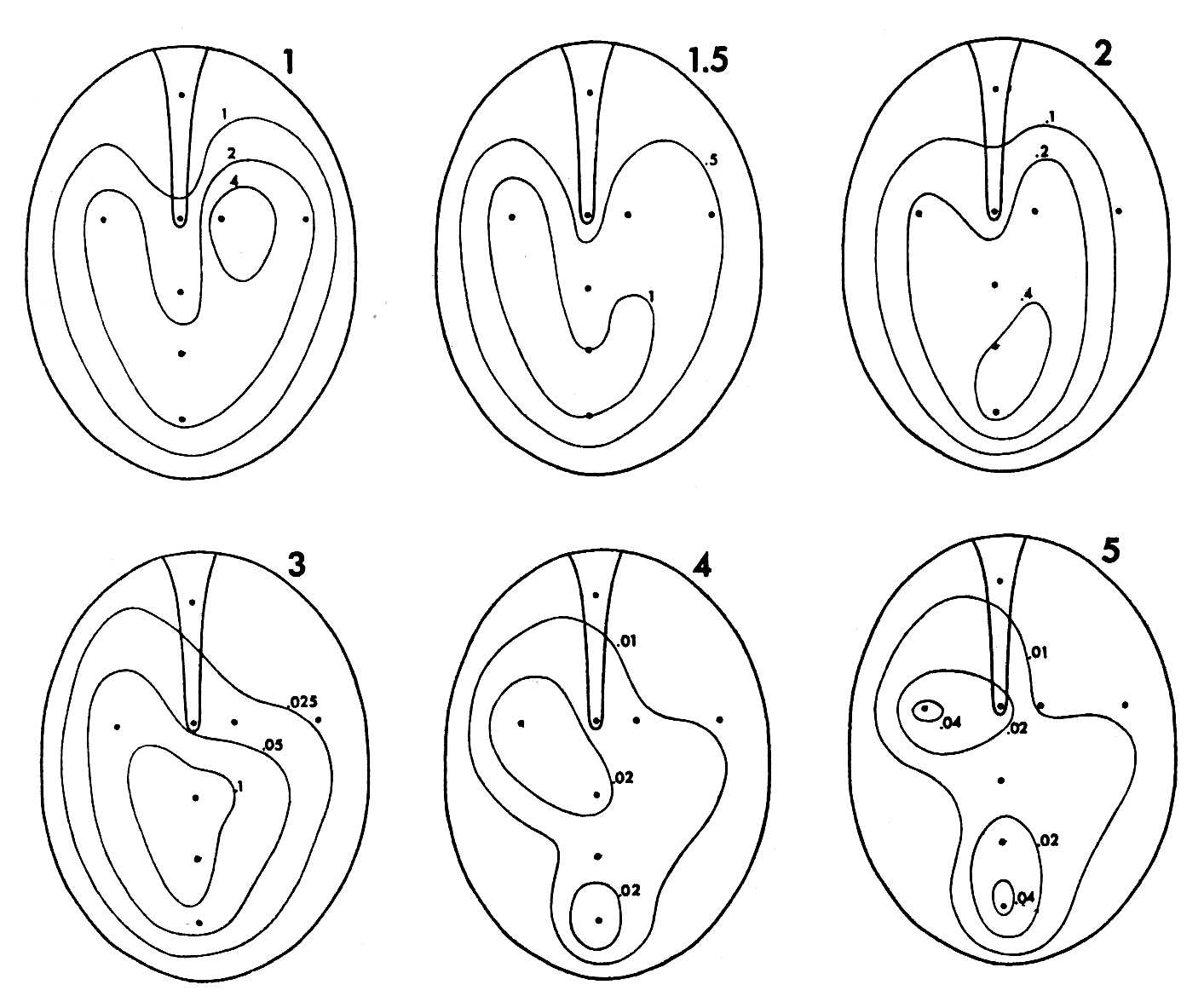
Color-coded vibration amplitude as a function of varying sound source direction and frequency of the signal.
Left: Internal coupling of the membranes intact. Right: One eardrum blocked. Because of the blocking, discrimination of the sound source direction is no longer possible!
(Measurements taken from Christensen-Dalsgaard 2006).
|
The next step in signal processing takes place on a neuronal level. An interesting phenomenon is the different treatment of low and high frequencies. This can be observed already at the stage of the basilar membrane and, as presented in the figures, it continues on the level of the neuronal network architecture. Here a network of exitatory and probably as well inhibitory neurons could be modeled and compared with experimental data.
This could hopefully complete our present knowledge about pressure-gradient receivers.
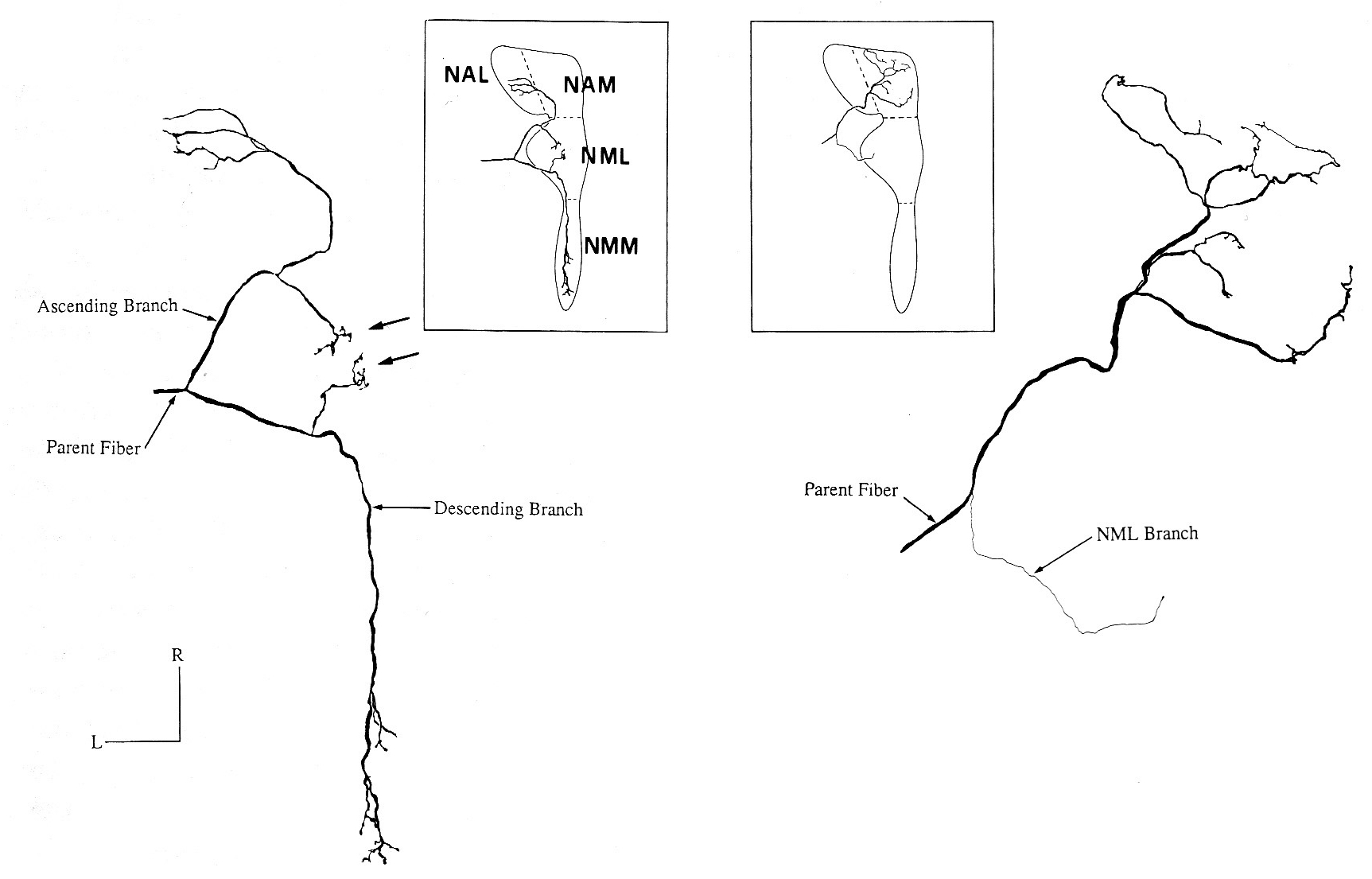
|
|
Different projections of low (left) and high (right) frequency fibers.
|
- Manley G.A., Köppl C., Johnstone B.M. Periphal auditory processing in the bobtail lizard Tiliqua rugosa - I to IV Journal of Computional Physiology 167:89-138 (1990)
- Manley G.A., Response patterns and peripheral origin of auditory nerve fibers in the monitor lizard, Varanus bengalensis Journal of Comparative Physiology A 118:249-260 (1977)
- Carr C.E., Christensen-Dalsgaard J. Evolution of the Auditory System in Reptiles and Birds Encyclopedia of Neuroscience, Springer, Berlin, 2006
- Christinsen-Dalsgaard J., Manley G.A. Directionality of the lizard ear Journal of Experimental Biology 208:1209-1217 (2005)
- Fletcher N.H. Acoustic Systems in Biology Oxford University Press, Oxford 1992

last modified 2007-11-05
by
webmaster@Franosch.org
|





